Revisión de fundamentos de la diferenciación celular: redes de TF, cromatina, rutas Wnt/Notch/Hedgehog y microambiente, con ejemplos y referencias canónicas clave.
Stem cell division and differentiation (Autor: User:Wykis · Licencia: Public domain · Fuente: Wikimedia Commons)
La diferenciación celular es un proceso dinámico que emerge de la interacción entre redes reguladoras de transcripción, el estado de la cromatina y la señalización intercelular, modulada por el microambiente. Este artículo sintetiza fundamentos moleculares de la diferenciación desde una perspectiva de base, orientando a estudiantes de grado y de posgrado en biología celular. Se trazan las relaciones entre la jerarquía de factores de transcripción, la memoria epigenética impartida por la cromatina y las rutas de señalización clásicas y contemporáneas, con ejemplos que conectan conceptos históricos con tecnologías modernas de análisis unicelular. En este marco, la idea de un paisaje epigenético, introducida como metáfora para entender destinos celulares, se complementa con hallazgos de reprogramación y diferenciación a nivel individual de células, enfatizando la importancia del contexto y la variabilidad biológica (Waddington, 1957).
La metáfora del paisaje epigenético describe la diferenciación como un recorrido por vallados y valles, en el que la topografía de la célula determina estados estables y rutas posibles de desarrollo.
En palabras de Waddington, la diferenciación no es un simple cambio lineal de estado, sino la interacción de redes reguladoras que guían a la célula hacia perfiles de expresión estables. Este marco se ha enriquecido con la demostración de que la identidad celular puede ser reconfigurada mediante la introducción de factores de transcripción y la manipulación de la cromatina, lo que llevó a la generación de células iPSC y a la exploración de trayectorias de línea celular a nivel único (Takahashi & Yamanaka, 2006). Paralelamente, los trabajos sobre la cromatina y las llamadas “pioneer factors” destacaron cómo ciertas proteínas pueden abrir regiones cerradas del genoma para permitir un programa transcripcional determinado (Zaret & Carroll, 2011).
Fundamentos conceptuales: redes de factores de transcripción y paisaje epigenético
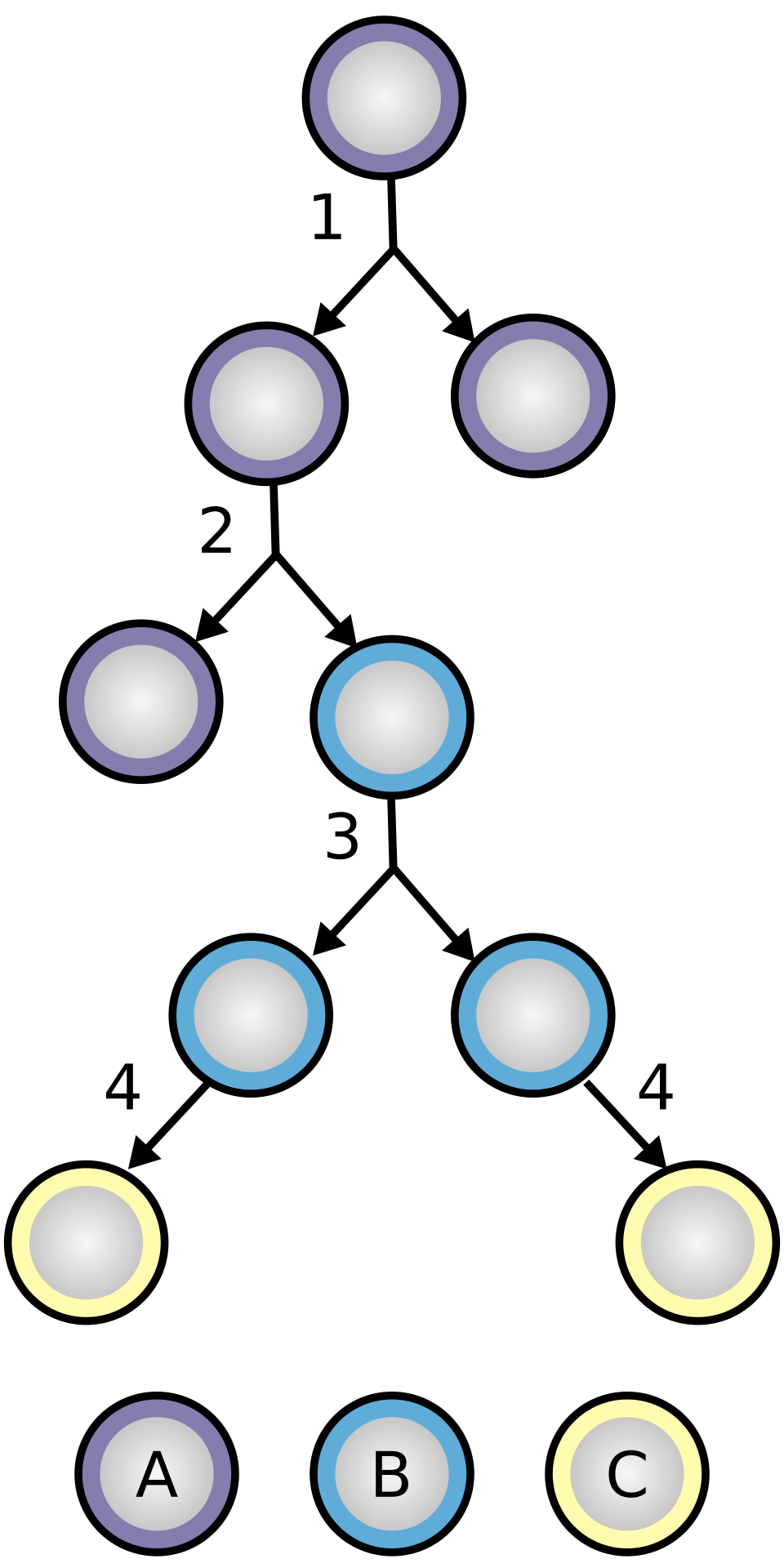
La diferenciación celular depende de redes de factores de transcripción (FT) que entrelazan regulación positiva y negativa para activar o reprimir programas gene expression. Estas redes no actúan aisladas: su dinámica está acoplada a la memoria epigenética que imparte la cromatina, determinando si un gen está en un estado accesible o silenciado. En etapas tempranas del desarrollo, las FT clave generan circuitos de auto-regulación y feed-forward loops que establecen estados de identidad. Cuando estas redes alcanzan un equilibrio inestable, pequeñas variaciones en el entorno o en la expresión de un factor pueden desviar la trayectoria hacia diferentes destinos celulares. Este concepto, que parte de la visión de paisaje de Waddington (1957), se ha apoyado en estudios de reprogramación y en análisis de transcriptómica de una sola célula, que revelan heterogeneidad y rutas preferenciales en la diferenciación (Treutlein et al., 2014).
La frontera entre estado de pluripotencia y diferenciación está mediada por cambios en la red de FT y por la remodelación de la cromatina. Los FT pueden funcionar como “pioneros” de la célula, estableciendo criptotipos que permiten o limitan la respuesta a señales externas. En la práctica, la diferenciación implica activar cascadas de factores que se refuerzan entre sí, y simultáneamente se cierran puertas genéticas mediante modificaciones de histonas y de la estructura de la cromatina. La interpretación de estas redes está influida por la memoria celular, que describe la persistencia de estados transcripcionales incluso cuando las señales han cambiado de manera transitoria (Takahashi & Yamanaka, 2006).
En un plano práctico, la conectividad entre TF y el estado cromatínico se estudia mediante enfoques de biología computacional y tecnologías de lectura del genoma. Por ejemplo, las regiones reguladoras que se abren durante la diferenciación pueden ser identificadas mediante cromatin accessibility assays, que marcan la acción de factores de transcripción y la prioridad de ciertos programas (Buenrostro et al., 2013). Esta perspectiva se integra con la noción de que los caminos de diferenciación no son lineales, sino que incluyen bifurcaciones y bucles de retroalimentación que pueden estabilizar o modificar la identidad de la célula.
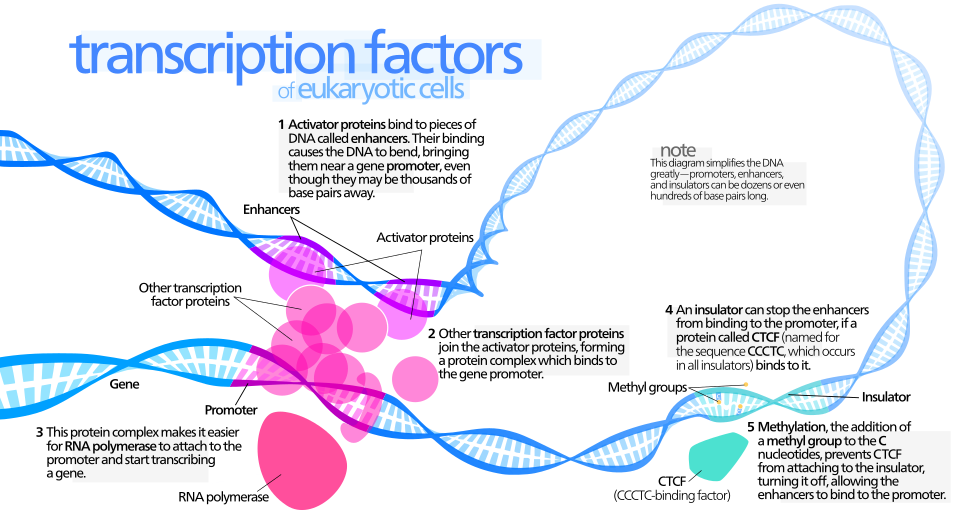
Arquitectura de la cromatina: accesibilidad, marcas y memoria celular Transcription Factors (Autor: Kelvin13 · Licencia: CC BY 3.0 · Fuente: Wikimedia Commons)
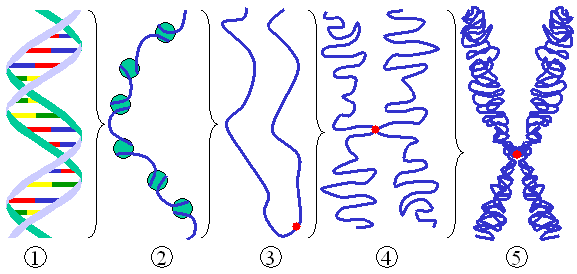
La cromatina regula en gran medida la capacidad de la maquinaria transcripcional para acceder a regiones génicas. Los cambios en la acetilación de histonas, la metilación de DNA y las modificaciones de las colas de histonas crean un paisaje de accesibilidad que facilita o restringe la expresión génica. En el contexto de la diferenciación, la remodelación de la cromatina no solo activa nuevos programas transcripcionales, sino que también consolida memorias celulares que se mantienen incluso ante cambios transitorios en el entorno. Técnicas como ATAC-seq han permitido mapear de manera global la accesibilidad cromatínica en poblaciones celulares y, más recientemente, a nivel de célula única, desentrañando la heterogeneidad de estados y la dinámica de apertura de regiones reguladoras en trayectorias de diferenciación (Buenrostro et al., 2013).
La memoria epigenética está también asociada a modificaciones de histonas y a la presencia de marcas que persisten a lo largo de divisiones celulares. Estas huellas permiten que una célula que experimenta un estímulo anterior conserve un “recuerdo” de su respuesta, modulando su respuesta futura y su capacidad de responder a señales similares. En este marco, el concepto de “pioneer factors” cobra especial relevancia: algunas proteínas pueden iniciar el acceso a regiones cerradas del genoma y facilitar la reprogramación de la identidad celular al establecer condiciones de competencia para otros factores (Zaret & Carroll, 2011).
Los factores pioneros no solo abren la cromatina, también establecen una ventana de tiempo en la cual la célula evalúa opciones de destino, integrando señales internas y externas para fijar la trayectoria de diferenciación.
La literatura moderna muestra que la cromatina y la regulación de la transcripción están íntimamente conectadas con la plasticidad celular: cambios sistemáticos en el estado de la cromatina acompañan la transición entre estados, y la memoria de dichos cambios soporta la persistencia de la identidad de la célula a través de divisiones y perturbaciones ambientales (Buenrostro et al., 2013; Zaret & Carroll, 2011).
Señalización clave en la diferenciación: Wnt, Notch y Hedgehog Chromatin chromosome (Autor: Magnus Manske · Licencia: CC BY-SA 3.0 · Fuente: Wikimedia Commons)
Las rutas de señalización extracelulares ejercen un control profundo sobre las decisiones de la célula durante la diferenciación, actuando como mediadores entre el microambiente y el programa genético interno. Las vías Wnt, Notch y Hedgehog son nodos centrales que coordinan la proliferación, la elección de destino y la compatibilidad de estados de diferenciación. Wnt regula la estabilidad de estados de progenitor y la especificación de linajes al modular la transcripción de genes diana en complejos de coactivación de la cromatina. Notch, por su parte, funciona como un interruptor dependiente del coentorno que puede mantener una población en un estado de progenitor o permitir la progresión hacia una identidad específica, dependiendo del equilibrio entre señales y la retroalimentación de la red. Hedgehog aporta señales de crecimiento y patrones de polaridad en diversos tejidos, interactuando con Wnt y Notch para afinar la respuesta frente a estímulos externos. La integración de estas rutas se observa en la regulación de programas transcripcionales que definen destinos celulares y ramificaciones de las trayectorias de diferenciación (Clevers & Nusse, 2012).
El reconocimiento de estas vías como módulos de control de la identidad celular se ha visto respaldado por estudios que combinan reprogramación, análisis de single-cell y manipulación de componentes de la cromatina. Por ejemplo, la activación coordinada de Wnt y Notch puede favorecer la adquisición de un estado de progenitor estable, mientras que la interrupción de una ruta puede favorecer rutas alternativas de diferenciación. A nivel experimental y terapéutico, comprender la lógica de estas vías facilita la disección de mecanismos de desarrollo y la optimización de estrategias de diferenciación dirigida, incluida la generación de linajes específicos a partir de células madre pluripotentes (Clevers & Nusse, 2012).
Microambiente y plasticidad celular
La identidad celular no está determinada únicamente por la red interna de transcription factors y el estado de la cromatina; el microambiente —incluyendo señales paracrinas, matriz extracelular, gradientes de oxígeno y interacciones célula-matriz— juega un papel decisivo en la plasticidad y la trayectoria de diferenciación. En contextos tisulares, las células enfrentan conjuntos complejos de señales que pueden converger o competir, generando estados intermedios o progresiones rápidas hacia destinos determinados. Este marco explica, en parte, por qué células idénticas pueden seguir trayectorias distintas cuando se exponen a condiciones distintas, y cómo los nichos celulares estabilizan o facilitan cambios de identidad (Treutlein et al., 2014).
La interacción entre el estado intrínseco de una célula y su entorno se ha observado en estudios de línea celular aislada y en tejidos complejos. Interpretar estas interacciones requiere aproximaciones que integren datos de expresión génica, chromatin state, y señales del microambiente, para predecir trayectorias de diferenciación y posibles estados intermedios. Las tecnologías de lectura de la actividad de genes y de cromatina a nivel único han permitido correlacionar cambios en el paisaje epigenético con cambios en el comportamiento de las células dentro de su nicho, subrayando la importancia de la heterogeneidad y la continuidad de los estados celulares (Treutlein et al., 2014; Buenrostro et al., 2013).
Tecnologías y enfoques para estudiar la diferenciación: desde el ensamblaje de redes hasta el análisis unicelular
La investigación moderna de diferenciación celular se apoya en un conjunto de tecnologías que permiten desentrañar redes de regulación y estados cromatínicos en poblaciones celulares y a nivel de una sola célula. El desarrollo de la transcriptómica de una célula, la epigenómica de alta resolución y las técnicas de lineage tracing han proporcionado herramientas para reconstruir jerarquías de linaje, rutas de diferenciación y la influencia de la historia celular en la respuesta a estímulos. El trabajo de Treutlein y coautores ilustra cómo los datos de expresión a nivel unicelular pueden identificar estados transitorios y rutas de reprogramación, revelando la heterogeneidad que subyace a procesos como la diferenciación y la conversión de estado (Treutlein et al., 2014).
En paralelo, la caracterización de la accesibilidad cromatínica y de las marcas de histonas ha permitido mapear los cambios estructurales que acompañan la diferenciación, facilitando la identificación de regiones reguladoras que definen la identidad de las células. Técnicas como ATAC-seq y ChIP-seq han sido fundamentales para entender cómo la topografía del genoma cambia durante la progresión de un linaje. Estos enfoques, combinados con modelos computacionales de redes de transcripción, proporcionan un marco para predecir trayectorias de destino y para diseñar estrategias de diferenciación hacia linajes deseados (Buenrostro et al., 2013; Zaret & Carroll, 2011).
La historia de la diferenciación también está escrita en la biología de la reprogramación. El hito de Yamanaka (2006) demostró que una combinación mínima de factores de transcripción puede reprogramar células adultas hacia un estado pluripotente, abriendo la puerta a la manipulación deliberada de programas de identidad celular y a la exploración de límites de plasticidad. Este logro ha impulsado la exploración de redes de TF y la interacción con la cromatina para entender mejor cómo se pueden compatibilizar o restringir rutas de diferenciación en contextos fisiológicos y experimentales (Takahashi & Yamanaka, 2006).
En conjunto, estas perspectivas integradas permiten una visión unificada: la diferenciación neuronal, hematopoyética, mesenquimal y de otros linajes resulta de la interacción entre una red de reguladores, un estado cromatínico dinámico y señales ambientales que guían o restringen la trayectoria. El conocimiento de estas interacciones facilita el desarrollo de modelos predictivos y la ingeniería de condiciones de cultivo que favorezcan la obtención de linajes específicos para investigación y medicina regenerativa (Treutlein et al., 2014; Buenrostro et al., 2013).
Perspectivas y retos actuales
A pesar de los avances, siguen existiendo retos considerables para entender plenamente la diferenciación celular. La naturaleza compleja de las redes de regulación, la heterogeneidad intrínseca de las células y la influencia del microambiente requieren enfoques multidisciplinares que integren biología molecular, biología computacional y física de sistemas. Las investigaciones que combinan datos de expresión génica a nivel de célula única con mapas de cromatina y de localización espacial permitirán construir modelos más precisos de cómo se decide el destino celular y cómo las perturbaciones pueden desviar trayectorias. En este ámbito, las referencias clásicas, como Waddington (1957) y los hitos de Yamanaka (2006), siguen guiando el marco conceptual; sin embargo, las pruebas empíricas se fortalecen a través de la generación de datasets de gran escala y la aplicación de métodos de lineage tracing que permiten reconstruir rutas de diferenciación con resolución sin precedentes (Treutlein et al., 2014).
En la práctica docente, este marco ofrece una base sólida para enseñar a estudiantes a interpretar datos experimentales: desde la identificación de redes de transcripción y módulos cromatínicos hasta la comprensión de cómo las señales extrínsecas condicionan la diferenciación y la memoria celular. La reflexión sobre estas dimensiones ayuda a formular preguntas de investigación coherentes y a diseñar experimentos que integren distintos niveles de regulación biológica para estudiar la diversidad de destinos celulares (Takahashi & Yamanaka, 2006; Buenrostro et al., 2013; Treutlein et al., 2014).
En síntesis, la diferenciación celular emerge como un proceso gobernado por una red de factores de transcripción acoplada a un paisaje cromatínico dinámico, modulada por rutas de señalización clave y modulada por el microambiente. Este marco conceptual facilita explicar cómo las células adquieren identidades estables y, al mismo tiempo, conservan capacidad de plasticidad frente a estímulos y perturbaciones, un equilibrio que constituye la base de la desarrollo, la homeostasis tisular y la regeneración.
Conclusión conceptual: la diferenciación celular es el resultado de una interacción entre redes de transcripción, cromatina y señales externas, donde la memoria epigenética y la historia de señalización determinan trayectorias de identidad, con una notable variabilidad a nivel celular individual que puede explorarse mediante enfoques de single-cell y multi-ómica.
Carmen Vidal Salgado
Analista de investigación científica especializada en biología celular y técnicas de imagen.
Carmen Vidal Salgado escribe sobre avances en biología celular, técnicas experimentales y bioinformática para una audiencia académica de habla hispana. Su trabajo se enfoca en sintetizar investigación reciente — desde microscopía de super-resolución hasta edición genética CRISPR — en formato accesible pero técnicamente riguroso. Las opiniones expresadas son interpretaciones del autor de la literatura científica publicada.